
 |
La parte más externa del núcleo
es la envuelta
nuclear.
|
ʮ Concepto: es el centro rector de toda la actividad celular.
Contiene el ADN codificando toda la información para la vida de la célula.
Controla todos los procesos de desarrollo. Es la característica esencial de las
células eucariotas con respecto a las células procariotas que no poseen un
núcleo definido. En el núcleo se realiza la duplicación y la transcripción del
ADN y el procesamiento de los ARN, formándose los tres tipos de ARN
principales, el ARN ribosómico, el ARN transferente y el ARN mensajero que
salen del núcleo y en el citoplasma es dónde se realiza la síntesis de
proteínas. La existencia de un núcleo implica que distintos procesos estén
separados. Si la célula está en división el aspecto del núcleo cambia.
® Historia:
El núcleo los vió Leeuwenhoek que dibujó eitrocitos de salmón
con núcleo.
En 1781 Fontana describió unos corpúsculos dentro de las
células epiteliales. Fue descrito con detalle en 1833 por Brown.
En el siglo XIX Waldeyer describió los cromosomas y ya en el
siglo XX Hershey y Chase identifican al ADN como la molécula donde se encuentra
contenida toda la información genética.
En 1953 Watson y Crick proponen la estructura de la doble
hélice para el ADN que fue el primer paso para el avance en el conocimiento de
los diferentes procesos en el campo de la información genética.
Características
generales:
A microscopía óptica si se tiñen las células con un colorante
denominado hematoxilina-eosina se observa el citoplasma teñido, se puede
observar un límite y en el centro se ve una estructura teñida de rojo intenso
con un pequeño corpúsculo más intenso.
Se puede ver el número
de núcleos por célula mediante la microscopía electrónica.
Los eritrocitos de los mamíferos y la plaquetas no tienen
núcleo.
Lo normal es que las células tengan un único núcleo pero por
ejemplo los hepatocitos del hígado tiene 2 núcleos, también algunas células de
la médula y de la corteza drenal y hay otras células multinluceadas como por
ejemplo los osteoclastos y también las células musculares estriadas
esqueléticas.
El tamaño varía de un tipo celular a otro, por término medio
se sitúa entre 5-15 m de diámetro. Siempre guarda relación con el tamaño del
citoplasma, de tal manera que hay una relación constante para cada tipo celular
(Vn / Vcit). Si esta constante cambia significa que la célula entra en división
y en las células hijas se vuelve a establecer esta misma relación.
La forma del núcleo se adapta a la forma de la célula en la
cual se encuentra. En las células esféricas el núcleo también es esférico, en
células cúbicas el núcleo también es esférico, en células aplanadas el núcleo
es alargado y en algunos casos es completamente irregular, por ejemplo en los
neutrófilos que son células esféricas el núcleo tiene una serie de lóbulos. La
forma puede cambiar.
Normalmente se sitúa en el centro de la célula pero en
ocasiones se encuentran desplazado. En los adipositos de grasa blanca que son
células cargadas con una gota de grasa, el núcleo se sitúa en la parte externa
de la célula. En las células musculares estriadas esqueléticas los núcleos se
sitúan en la periferia porque en el centro se disponen los componentes del
citoesqueleto. En las células especializadas en la secreción, el núcleo se
sitúa en el tercio inferior de la célula.
Composición
química:
- Envuelta
nuclear: el 75% son proteínas y el 25% restante son lípidos. En una membrana
muy activa.
- Interior
nuclear: principalmente es agua, si se considera el peso en seco hay un 15% de
ADN, un 5% de ARN y el resto son proteínas aunque también hay sales minerales,
iones, etc. Dentro de esas proteínas las hay de diferentes tipos.
a) Proteínas
enzimáticas: como la ADN y la ARN polimerasa.
b) Proteínas no
enzimáticas: las hay básicas que son las histonas que se dividen en 5 tipos
diferentes (H1, H2A, H2B, H3 y H4) y que son las que se unen al ADN y lo
empaquetan. Y también las hay no básicas que son las que intervienen en la
transcripción del ADN en la estructuración de los cromosomas.
® Ultraestructura:
La parte más externa del núcleo es la envuelta nuclear.
La envuelta nuclear está constituida por dos membranas
(membrana nuclear interna y membrana nuclear externa) que presentan las
características de todas las membranas biológicas separadas las dos membranas
por el espacio perinuclear.
En la membrana nuclear externa están pegados ribosomas,
mirando hacia el citoplasma, y se puede ver que la envuelta nuclear es continua
con las cisternas del RER. En algunos casos de tal manera que el espacio
perinuclear es continuo con la luz de las cisternas del RE.
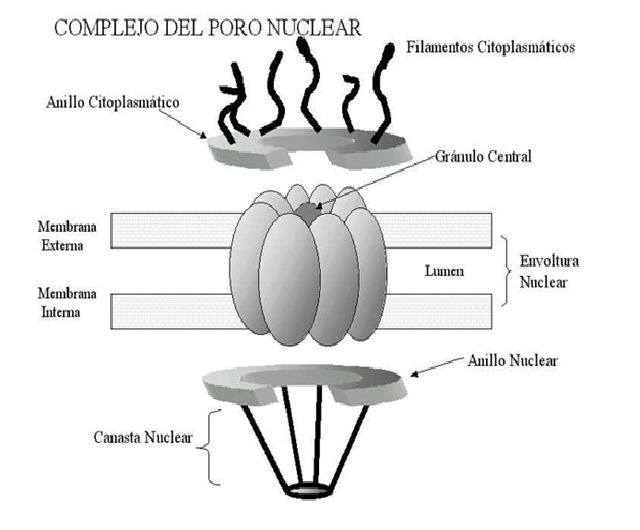
La envuelta nuclear no es continua, en ciertos puntos está
interrumpida. Esos puntos se denominan poros nucleares, donde se sitúan unos
grandes complejos proteicos que en conjunto a todo el complejo se le conoce con
el nombre de complejo de poro nuclear.
Hacia el interior del núcleo y pegado a la membrana nuclear
interna se encuentra la lámina nuclear constituida por proteínas de filamentos
intermedios. En el resto del núcleo hay cromatina que puede aparecer en dos
formas: Eucromatina (es la cromatina más condensada) y heterocromatina (es la
cromatina menos condensada y que a su vez puede aparecer en dos formas
distintas, constitutiva y facultativa).
La heterocromatina suele aparecer por dentro de la lámina
nuclear, en la parte externa del núcleo, también rodeando al nucleolo y como
acúmulos dispersos dentro del núcleo.
También se encuentra el nucleolo que es una región del núcleo
donde se forman los ribosomas y tanto la cromatina como el nucleolo se
encuentran en un medio líquido llamado nucleoplasma o carioplasma.
Envuelta nuclear:
Es la estructura que separa el contenido del núcleo del
contenido del citoplasma y actúa como una barrera selectiva que impide el paso
libre de moléculas entre el exterior y el interior del núcleo. Controla el paso
de sustancias entre el núcleo y el citoplasma.
Ese intercambio de sustancias entre el núcleo y el citoplasma
se realiza a través de los poros nucleares. Se realiza el paso selectivo de
proteínas y de ARN; por tanto, los complejos de poro van a tener una función en
el control de la expresión génica.
Está constituida por dos membranas que son igual que las
demás membranas biológica,s asimétricas, etc. Tienen un espesor de alrededor de
unos 7 nm y el espacio que dejan entre ellas es de 25-40 nm.
La membrana nuclear externa tiene receptores para la unión de
ribosomas. Los ribosomas realizan la síntesis de proteínas y esas proteínas
pasan al espacio perinuclear; por tanto, el espacio perinuclear es muy similar
en cuanto a su composición.
Es la estructura que separa el contenido del núcleo del
contenido del citoplasma y actúa como una barrera selectiva que impide el paso
libre de moléculas entre el exterior y el interior del núcleo. Controla el paso
de sustancias entre el núcleo y el citoplasma.
Ese intercambio de sustancias entre el núcleo y el citoplasma
se realiza a través de los poros nucleares. Se realiza el paso selectivo de
proteínas y de ARN; por tanto, los complejos de poro van a tener una función en
el control de la expresión génica.
Está constituida por dos membranas que son igual que las
demás membranas biológica,s asimétricas, etc. Tienen un espesor de alrededor de
unos 7 nm y el espacio que dejan entre ellas es de 25-40 nm.
La membrana nuclear externa tiene receptores para la unión de
ribosomas. Los ribosomas realizan la síntesis de proteínas y esas proteínas
pasan al espacio perinuclear; por tanto, el espacio perinuclear es muy similar
en cuanto a su composición.
La membrana nuclear interna contiene un receptor para unirse
a la lámina nuclear.
Lo más característica de la envuelta nuclear son los poros
nucleares.
A nivel del poro nuclear la membrana nuclear interna y
externa se fusionan. Los poros nucleares aparecen en el núcleo de todas las
células.
Por término medio en las células de levaduras hay unos 200
poros por núcleo, en células humana en proliferación hay entre 3000 y 5000
poros nucleares por núcleo, en los ovocitos puede haber hasta 50000000 de poros
nucleares por núcleo. El núcleo de los espermatozoides no presenta poros
nucleares.
El poro nuclear es una estructura muy grande. Tiene un
diámetro de aproximadamente 120 nm, pero el agujero real es mucho más pequeño,
tiene entre 45-50 nm y se debe a que los poros nucleares se disponen en un
complejo de proteínas que se conocen como nucleoporinas.
En las células humanas se han encontrado entre 50 y 100
nucleoporinas distintas formando parte del complejo de poro nuclear.
}
Las proteínas que forman parte del complejo de Golgi se
disponen siguiendo una disposición octamérica. Estas nucleoporinas se componen
de:
- Complejo
anular-radial: es un grupo de 8 proteínas que se disponen pegadas a las
membranas del núcleo. Se ancla a la membrana nuclear externa y a la membrana
nuclear interna y hace que esas dos membranas sean distintas. En el centro se
dispone otro complejo proteico denominado complejo transportador.
- Complejo
nucleoplásmico: en el lado del nucleoplasma.
Las sustancias apolares pequeñas son las sustancias que
pueden pasar a través de las membranas nucleares sin ayuda de los poros
nucleares.
A través del complejo de poro hay tres tipos de transporte
distinto:
- Difusión
pasiva: las sustancias que pasan a través del complejo de poro no necesitan
interaccionar con las nucleoporinas. Es un transporte rápido pero sólo pasan
moléculas pequeñas que tengan un bajo peso molecular y parece que el tamaño de
las partículas tiene que ser menor de 9 nm que sería el tamaño del poro real
que deja el complejo transportador.
- Difusión
facilitada: permite el paso de moléculas de mayor tamaño. Es necesario que las
moléculas a transportar establezcan interacciones con las nucleoporinas de tal
manera que se modifica la conformación del complejo de poro.
- Transporte
activo: para el transporte activo son necesarias proteínas transportadoras que
establezcan interacciones con los nucleoporinas que se denominan
transportadores nucleares. Hay diferentes tipos de proteínas transportadores,
pueden ser importinas (las que llevan las sustancias hacia el núcleo. Reconocen
señales de localización nuclear (NLS)), exportinas (las que sacan las
sustancias desde el núcleo al citoplasma. Reconocen señales de localización plasmática)
y las transportinas (que permiten el paso de en dos direcciones, del núcleo al
citoplasma y del citoplasma al núcleo. Reconocen señales lanzadera (NS)).
El transporte activo está mediado por las proteínas Ran que
están unidas a GTP o a GDP, que media la unión del transportador y de la
molécula a transportar (ran-GTP y ran-GDP). Conllevan gasto de energía en forma
de GTP.
Laminillas
anilladas: en relación con la envuelta nuclear. Aparecen en el citoplasma de
algunos tipos celulares (células en mitosis, embrionarias y células tumorales).
Tienen las mismas características que la envuelta nuclear
(con complejo de poro).
Se supone que son reservorios del complejo de poro que se
integrarían en la envuelta nuclear en momentos en que el incremento en la tasa
de transporte fuese incrementado. No tienen lámina nuclear.
El espesor de las membranas es de alrededor de 7 nm y el
espacio que queda entre ellas es del orden de entre 30-50 nm.
Lámina nuclear. Es
una red de proteínas de entre 30-100 nm de espesor que se sitúa por dentro de
la membrana nuclear interna.
Está constituida por filamentos intermedios de 10 nm de
diámetro.
Los filamentos de la lámina nuclear están constituidos por 3
proteínas distintas:
- La lámina o
laminina A.
- La lámina o
laminina B.
- La lámina o
laminina C.
La lámina B se une a la membrana nuclear interna, esa
membrana nuclear interna tiene un receptor para la lámina B. A su vez la A y la
C se unen a la B por un lado y por otro lado se unen a la cromatina. Es un
sistema para mantener la cromatina dentro del núcleo.
Puede ensamblarse y desensamblarse, cuando la célula entre en
división la lámina nuclear se desorganiza porque se produce la desfosforilacion
de las proteínas de las láminas. Sin embargo, al final de la mitosis tiene
lugar el proceso contrario, las proteínas se fosforilan.
Esa lámina nuclear es la responsable del ensamblaje y
desensamblaje de la envuelta nuclear durante los procesos de división celular.
Los filamentos de la lámina nuclear están constituidos por 3
proteínas distintas:
- La lámina o
laminina A.
- La lámina o
laminina B.
- La lámina o
laminina C.
La lámina B se une a la membrana nuclear interna, esa membrana
nuclear interna tiene un receptor para la lámina B. A su vez la A y la C se
unen a la B por un lado y por otro lado se unen a la cromatina. Es un sistema
para mantener la cromatina dentro del núcleo.
Puede ensamblarse y desensamblarse, cuando la célula entre en
división la lámina nuclear se desorganiza porque se produce la desfosforilacion
de las proteínas de las láminas. Sin embargo, al final de la mitosis tiene
lugar el proceso contrario, las proteínas se fosforilan.
Esa lámina nuclear es la responsable del ensamblaje y
desensamblaje de la envuelta nuclear durante los procesos de división celular.
Nucleoplasma: es
el medio líquido en el que está la cromatina y el nucleolo. Es una fase acuosa
que contiene fundamentalmente proteínas (proteínas relacionadas con el
metabolismo de ácido nucleicos, proteínas que intervienen en la regulación de
la expresión génica (factores de trascripción, moduladores, represores, etc)).
También hay cofactores, minerales, iones (magnesio y calcio
sobre todo, también aparece sodio, potasio, etc) NAD, ATP y además otras
moléculas.
Además de la lámina nuclear se discute si hay un esqueleto de
proteínas similar al citoesqueleto del citoplasma. Lo que sí se sabe es que
existe actina dentro del núcleo y tubulina tanto alfa como beta tubulina pero
algo distintas a las citoplasmáticas.
También se han encontrado proteínas no identificadas que unen
entre sí determinadas secuencias del ADN llamadas MAR o SAR (son regiones
asociadas a la matriz).
Toda esa matriz de proteínas organiza los plegamientos de la
cromatina y de esa manera regula la replicación y la transcripción del ADN.
Dentro del núcleo hay cromatina y está el nucleolo.
Cromatina: está
constituida principalmente por ADN y por proteínas y, además, puede aparecer
con aspectos distintos. Toda esa cromatina es ADN y proteínas.
, moduladores, represores, etc)).
También hay cofactores, minerales, iones (magnesio y calcio
sobre todo, también aparece sodio, potasio, etc) NAD, ATP y además otras
moléculas.
Además de la lámina nuclear se discute si hay un esqueleto de
proteínas similar al citoesqueleto del citoplasma. Lo que sí se sabe es que
existe actina dentro del núcleo y tubulina tanto alfa como beta tubulina pero
algo distintas a las citoplasmáticas.
También se han encontrado proteínas no identificadas que unen
entre sí determinadas secuencias del ADN llamadas MAR o SAR (son regiones
asociadas a la matriz).
Toda esa matriz de proteínas organiza los plegamientos de la
cromatina y de esa manera regula la replicación y la transcripción del ADN.
Dentro del núcleo hay cromatina y está el nucleolo.
Cromatina: está
constituida principalmente por ADN y por proteínas y, además, puede aparecer
con aspectos distintos. Toda esa cromatina es ADN y proteínas.
Proteínas que se
asocian al ADN:
- Histonas: son
proteínas básicas. Se recambian muy poco en la célula, únicamente en la fase G1
y S del ciclo celular.
a) Nucleosómicas:
son la H2A, H2B, H3 y H4. Las cuatro forman los nucleosomas. Cada uno de esos
nucleosomas es un disco de un diámetro de alrededor de 11 nm que contiene dos
copias de cada una de estas histonas. Son proteínas básicas con restos de
lisina y arginina.
b) No
nucleosómicas: es la H1. No forman parte de los nucleosomas. Es mayor que las
otras proteínas histonas y es bastante heterogénea. Tiene una región globular y
dos colas en los extremos C- y N- terminal.
- No histonas:
la mayoría son ácidas del tipo de fosfoproteínas. También se asocian al ADN.
Tiene una velocidad de recambio muy alta. Se sintetizan a lo largo de todo el
ciclo celular. Las más conocidas son:
a) Nucleoplasmina:
que se unen a las histonas H2A y H2B.
b) Proteína N1:
que se une a las histonas H3 y H4.
c) PCNA (antígeno
nuclear de proliferación) Se desplaza a lo largo de la fibra de ADN. Parece que
intervienen en la replicación, reparación y recombinación del ADN. Actúa como
una proteína unida a la ADN polimerasa y se relaciona con proteínas que
intervienen en la control del ciclo celular. Aparecen en células que están en
división. Se utiliza como marcador de células proliferativas.
d) Enzimas que
intervienen en la duplicación del ADN.
En algunos tipos celulares no hay histonas. En los
espermatozoides de algunas especies las histonas son sustituidas por otras
proteínas de menor peso molecular que son las protaminas para conseguir un
mayor empaquetamiento del ADN.
Genoma: es toda la
información génica contenida en la cromatina. Es variable de una especie a
otra.
El genoma de eucariotas es mayor que el de procariotas.
Los anfibios tienen mayor peso génico que las aves y puede
haber también diferencia en cuanto al sexo. En un organismo diploide el genoma
está replicado dos veces.
En los seres humanos el genoma se encuentra duplicado.
Tenemos 22 pares de cromosomas que son los autosomas y los 2 cromosomas
sexuales. Tenemos 46 moléculas de ADN. Cada cromosoma es una molécula de ADN.
El genoma se divide en dos clases:
- Genoma
codificante.
- Genoma no
codificante.
El 40% del ADN son porciones no codificantes. Se alternan
porciones codificantes y porciones no codificantes, de diferentes tamaños. Las
porciones codificantes se denominan gen. Cada región de ADN que produce una
molécula de ARN funcional es un gen y dentro de las moléculas de ADN se
encuentran genes y porciones no codificantes.
Dentro de un gen se encuentran diferentes porciones. Los
genes normalmente llevan secuencias reguladoras de la expresión del gen. En el
gen se encuentran alternando intrones y exones que es la porción del ADN que se
transcribe al ARN.
El genoma se divide en dos clases:
- Genoma
codificante.
- Genoma no
codificante.
El 40% del ADN son porciones no codificantes. Se alternan
porciones codificantes y porciones no codificantes, de diferentes tamaños. Las
porciones codificantes se denominan gen. Cada región de ADN que produce una
molécula de ARN funcional es un gen y dentro de las moléculas de ADN se
encuentran genes y porciones no codificantes.
Dentro de un gen se encuentran diferentes porciones. Los
genes normalmente llevan secuencias reguladoras de la expresión del gen. En el
gen se encuentran alternando intrones y exones que es la porción del ADN que se
transcribe al ARN.
Al ARN se transcriben tanto los intrones como los exones pero
no es un ARN funcional, sino que tiene que producirse una serie de procesos de
maduración del ARN o “splicing” y se elimina la porción de los intrones
quedando los exones y la molécula de ARN ya es funcional.
Las secuencias reguladoras del ADN no se transcriben, sólo se
replican. En ocasiones se necesitan varias copias de un gen para sintetizar
muchas proteínas.
El conjunto de gen que codifica para un mismo ARN para una
misma proteína se denomina familia de genes. Dentro de la familia hay pequeños
cambios.
En otros casos se pueden transcribir diferentes miembros de
una familia de genes en diferentes tejidos o en diferentes fases de desarrollo.
Estos procesos se han consolidado a lo largo de la evolución.
Los pseudogenes son genes que no son funcionales. En el caso
de la hemoglobina se conoce que hay 2 genes que no son funcionales por
mutaciones.
Parte del ADN no codificante se corresponde con el genoma no
codificante que constituye el 40% de todo el ADN. Consiste en secuencias
repetitivas algunas de las cuales se repiten millones de veces. No se conoce
bien su función pero podrían ser secuencias reguladoras de la expresión de los
genes o podría tener función estructural (Ej. Unión a la lámina nuclear).
Hay dos tipos de ADN no codificante:
- ADN satélite:
contiene formaciones en tandem (repetidas una a continuación de otras). Tienen
entre 5 y 200 nucleótidos y se repiten millones de veces por genoma, pero
siempre una a continuación de otra, representan alrededor de entre el 10y el
20% de todo el ADN. No aportan ninguna información genética funcional. Algunas
sí empeñan funciones importantes en la estructura de los cromosomas. Son
pequeñas secuencias repetidas una a continuación de otra. Hay también
minisatélites que son del orden de 15 pares de bases que se repiten miles de
veces y microsatélites que son del orden de 5 pares de bases repetidas cientos
de veces, pero siempre en tandem.
- ADN disperso:
son secuencias de ADN que se repiten pero dispersas en el genoma. Hay a su vez
dos tipos:
a) Secuencias
“SINE”: son elementos cortos dispersos y de alrededor de 300 pares de bases y
hay alrededor de un millón de ellas dispersas en el genoma. Representan
alrededor del 10% del genoma. Estas secuencias se transcriben a ARN pero ese
ARN no codifica proteínas.
b) Secuencias
“LINE”: pueden tener 6000 pares de bases y se repiten del orden de 50000 veces.
Tienen entre 5 y 200 nucleótidos y se repiten millones de
veces por genoma, pero siempre una a continuación de otra, representan
alrededor de entre el 10y el 20% de todo el ADN. No aportan ninguna información
genética funcional. Algunas sí empeñan funciones importantes en la estructura
de los cromosomas. Son pequeñas secuencias repetidas una a continuación de
otra. Hay también minisatélites que son del orden de 15 pares de bases que se
repiten miles de veces y microsatélites que son del orden de 5 pares de bases
repetidas cientos de veces, pero siempre en tandem.
- ADN disperso:
son secuencias de ADN que se repiten pero dispersas en el genoma. Hay a su vez
dos tipos:
a) Secuencias
“SINE”: son elementos cortos dispersos y de alrededor de 300 pares de bases y
hay alrededor de un millón de ellas dispersas en el genoma. Representan
alrededor del 10% del genoma. Estas secuencias se transcriben a ARN pero ese
ARN no codifica proteínas.
b) Secuencias
“LINE”: pueden tener 6000 pares de bases y se repiten del orden de 50000 veces.
® Condiciones
necesarias para que el ADN constituya un cromosoma: Se necesita tener una
secuencia de ADN obligatoria que son importantes porque permiten que el
cromosoma se puede duplicar. Si el cromosoma no se puede duplicar no sería un
cromosoma.
- Orígenes de
replicación (ORI): para que el ADN se pueda duplicar. Hay varias a lo largo de
un cromosoma. Sin ellos no se podría duplicar el ADN.
- Centrómeros:
son secuencias de ADN a las cuales se pueden unir proteínas pero unas proteínas
particulares, las proteínas del cinetocoro. Sin centrómeros no se puede dividir
correctamente la célula y no se tienen cromosomas. En los seres humanos son secuencias satélite, secuencias muy
repetidas y en tandem.
- Telómeros:
son secuencias repetidas en tandem de ADN situadas en los extremos de los
cromosomas. Desempeñan un papel fundamental en la replicación y en el
mantenimiento del cromosoma. En los seres humanos esa secuencia se conoce y es
AGGGTT y se repite entre cientos y miles de veces. Esas secuencias hacen que en
los extremos de los cromosomas se formen unos lazos para proteger los extremos
del ADN. La telomelina permite que se
conserven los extremos de los cromosomas. El mantenimiento de estos Telómeros parece
que está relacionado con el envejecimiento, se pierde parte de los extremos de
los telómeros.
® El ADN se
empaqueta en los nucleosomas. El ADN se enrolla alrededor de los nucleosomas
que están constituidos por 8 histonas.
Cada 200 pares de bases aparece un nucleosoma. El ADN da 1,6 vueltas alrededor de esas
proteínas histonas. La fibra nucleosómica es el ADN enrollado en los
nucleosomas.
Pero en el empaquetamiento también interviene la histona H1
que puede que se una por una parte al nucleosoma y; por otra parte, al ADN
ayudando a empaquetar el ADN.
La estructura que se forma por la unión entre el nucleosoma
con la fibra de ADN a su alrededor y la histona H1 se denomina cromatosoma.
Este empaquetamiento da una fibra de un diámetro de 11 nm.
El ADN son unidades repetidas de cromatosomas.
Las secuencias que se están transcribiendo se supone que no
están en los nucleosomas y la presencia de los nucleosomas entre el ADN parece
que está determinada por dos factores. En primer lugar parece que los nucleosomas
están en zonas ricas en pares de bases A-T porque parece que esas regiones son
más fáciles de “doblar”. Y también parece que viene determinado por la
presencia de otras proteínas que normalmente son proteínas de regulación
génica.
La fibra de 11 nm no queda así sino en forma de fibras más
empaquetadas (con un diámetro de alrededor de 30 nm).
® El ADN se
empaqueta en los nucleosomas. El ADN se enrolla alrededor de los nucleosomas
que están constituidos por 8 histonas.
Cada 200 pares de bases aparece un nucleosoma. El ADN da 1,6 vueltas alrededor de esas
proteínas histonas. La fibra nucleosómica es el ADN enrollado en los
nucleosomas.
Pero en el empaquetamiento también interviene la histona H1
que puede que se una por una parte al nucleosoma y; por otra parte, al ADN
ayudando a empaquetar el ADN.
La estructura que se forma por la unión entre el nucleosoma
con la fibra de ADN a su alrededor y la histona H1 se denomina cromatosoma.
Este empaquetamiento da una fibra de un diámetro de 11 nm.
El ADN son unidades repetidas de cromatosomas.
Las secuencias que se están transcribiendo se supone que no
están en los nucleosomas y la presencia de los nucleosomas entre el ADN parece
que está determinada por dos factores. En primer lugar parece que los nucleosomas
están en zonas ricas en pares de bases A-T porque parece que esas regiones son
más fáciles de “doblar”. Y también parece que viene determinado por la
presencia de otras proteínas que normalmente son proteínas de regulación
génica.
La fibra de 11 nm no queda así sino en forma de fibras más
empaquetadas (con un diámetro de alrededor de 30 nm).
Se supone que esa fibra de 30 nm es la fibra cromatosómica
mucho más enrollada. También hay intervalos con porciones de ADN que no están
empaquetadas que ahí es donde se unen proteínas específicas de unión al ADN.
Esa fibra de 30 nm se puede empaquetar más dando fibras de
alrededor de 300 nm.
La fibra de 300 nm da otra agrupación de 700 nm y la de 700
nm da ya los cromosomas.
En el caso de los cromosomas se piensa que tiene un esqueleto
proteico y alrededor se enrolla el ADN.
El diámetro de los cromosomas es del orden de 1400 nm y sólo
son observables cuando la célula está en división junto con el de 700 nm. El
resto sólo los vemos cuando la célula se encuentra en interfase.
La eucromatina es la fibra de 11 nm que es la que se puede
transcribir pero a partir de ahí hay diferentes opiniones, algunos autores
piensan que a partir de 30 nm es heterocromatina aunque para otros es a partir
de la fibra de 300 nm, que la Eucromatina sería la de 11 nm y la de 30 nm.
Se presentan diferencias entre la eucromatina y la
heterocromatina que es la cromatina inactiva, que es la que no se transcribe,
el ADN presenta metilaciones y las histonas no acetiladas; sin embargo, en la
eucromatina el ADN no está metilado y las histonas están acetiladas.
Dentro de la heterocromatina se distinguen dos tipos:
- Heterocromatina
constitutiva: es aquella que nunca se transcribe como por ejemplo secuencias de
los centrómeros y de los telómeros.
- Heterocromatina
facultativa: es aquella que no se transcribe en un tipo celular pero sí se
transcribe en otros tipos celulares. En otro tipo celular puede aparecer como
eucromatina.
El proceso de inactivación de una parte de un ADN es lo que
sucede con el cromosoma X en el caso de las hembras. Uno de los cromosomas X de
las hembras se inactiva y aparece como heterocromatina y se inactiva en etapas
primarias del desarrollo.
A microscopía electrónica gracias a los neutrófilos se puede
distinguir si esa sangre es de mujer o de hombre.
Los cromosomas ocupan posiciones determinadas dentro del
núcleo.
Normalmente los centriolos y los telómeros se disponen en la
parte externa del núcleo y además dentro de cada par de cromosomas esos
cromosomas ocupan sitios distintos dentro del núcleo. Esos lugares se denominan
dominios cromosómicos y entre esos dominios cromosómicos se encuentran unos
espacios denominados dominios intercromosómicos. Parece que esos dominios
intercromosómicos son zonas libres de ADN a donde van pasando los ARN que se
están transcribiendo a partir del ADN. Las zonas que se transcriben del cromosoma
sería la zona más externa de los dominios cromosómicos.
transcribe en otros tipos celulares. En otro tipo celular
puede aparecer como eucromatina.
El proceso de inactivación de una parte de un ADN es lo que
sucede con el cromosoma X en el caso de las hembras. Uno de los cromosomas X de
las hembras se inactiva y aparece como heterocromatina y se inactiva en etapas
primarias del desarrollo.
A microscopía electrónica gracias a los neutrófilos se puede
distinguir si esa sangre es de mujer o de hombre.
Los cromosomas ocupan posiciones determinadas dentro del
núcleo.
Normalmente los centriolos y los telómeros se disponen en la
parte externa del núcleo y además dentro de cada par de cromosomas esos
cromosomas ocupan sitios distintos dentro del núcleo. Esos lugares se denominan
dominios cromosómicos y entre esos dominios cromosómicos se encuentran unos
espacios denominados dominios intercromosómicos. Parece que esos dominios
intercromosómicos son zonas libres de ADN a donde van pasando los ARN que se
están transcribiendo a partir del ADN. Las zonas que se transcriben del
cromosoma sería la zona más externa de los dominios cromosómicos.
Dentro del núcleo existen unas regiones denominadas dominios
funcionales dentro del núcleo. Los distintos procesos que se están realizando
dentro del núcleo se realizarían esos procesos en zonas determinadas del núcleo
de tal manera que la organización más interna del núcleo sería un reflejo de
todos los procesos realizados en el núcleo.
® Dominios
funcionales dentro del núcleo:
- Fábricas de
replicación o replicones: zonas donde tiene lugar la replicación del ADN. La
bromodeoxiuridina se puede incorporar a las células y con esta sustancia se
puede luego observar los sitios donde se está replicando el ADN.
- Las motas
nucleares o llamadas puntos de Splicing: es dónde se produce la maduración de
los ARN. Se han encontrado unos complejos de ARN y proteínas que son los
responsables de llevar a cabo la maduración de los ARN. Esos ARNs de denominan
ARNsn (ARN nuclear pequeño). Se han descrito 5 tipos distintos de esos ARNsn en
esas zonas que se unen a proteínas (se han descrito hasta ahora 6 proteínas
distintas de unión a esos ARN) y ese conjunto forman partículas ribonucleicas
que se conocen como espliceosomas que se pueden detectar al microscopio.
- Cuerpos
nucleares: son zonas que aparecen como diferentes dentro del núcleo. Se
distinguen dos tipos, cuerpos espirales y cuerpos PML. En la zona de los
cuerpos espirales se han visto partículas constituidas por ARNsn y por
proteínas y se supone que son los sitios donde se ensamblan esas partículas de
proteínas con ARNsn. Los cuerpos PML se desconoce su función, no aparecen ARNsn
unidos a proteínas parece que no están relacionados ni con la transcripción ni
con la replicación.
- Nucleolo:
donde se lleva a cabo la síntesis de los ARNr y la unión de los ARNr a
proteínas. Es el único dominio funcional dentro del núcleo y se puede
distinguir a microscopía electrónica. Se compone de diferentes partes.
® Nucleolo: el
nucleolo es una región que se organiza alrededor de la región de los cromosomas
donde están los genes que codifican los ARNr. Las regiones de los cromosomas
donde se encuentran los genes que codifican para los ARNr es lo que se conoce
como organizadores nucleolares. Esos genes en la especie humana se localizan en
5 cromosomas (en el cromosoma 13, 14, 15, 21 y 22). Además son genes que están
repetidos uno a continuación de otro, en tandem. Hay 10 organizadores partículas
constituidas por ARNsn y por proteínas y se supone que son los sitios donde se
ensamblan esas partículas de proteínas con ARNsn.
Los cuerpos PML se desconoce
su función, no aparecen ARNsn unidos a proteínas parece que no están
relacionados ni con la transcripción ni con la replicación.
- Nucleolo:
donde se lleva a cabo la síntesis de los ARNr y la unión de los ARNr a
proteínas. Es el único dominio funcional dentro del núcleo y se puede
distinguir a microscopía electrónica. Se compone de diferentes partes.
® Nucleolo: el
nucleolo es una región que se organiza alrededor de la región de los cromosomas
donde están los genes que codifican los ARNr. Las regiones de los cromosomas
donde se encuentran los genes que codifican para los ARNr es lo que se conoce
como organizadores nucleolares. Esos genes en la especie humana se localizan en
5 cromosomas (en el cromosoma 13, 14, 15, 21 y 22).
Además son genes que están
repetidos uno a continuación de otro, en tandem. Hay 10 organizadores
nucleolares. EN el nucleolo es donde se codifican los ARNm de 28 s, 18s y 5,8s
y el de 5s se sintetiza fuera del nucleolo y después ya se ven los ARNr que se
sintetizan en el nucleolo.
A partir del ADN que contienen los genes que codifica para
ARNr se sintetiza un preARNr de 45s que todavía no es funcional. Se produce un
proceso de maduración que se rompe y da 3 porciones distintas, una de 18 s, 28
s y 5,8 s y a esos también se une el ARNr de 5 s que se ha sintetizado fuera
del nucleolo. La subunidad menor está constituida por ARNr de 18 s más
proteínas y la subunidad mayor está constituido por ARNr de 28 s, 5,8 s y 5s.
Los RNAsno (ARN nucleares pequeños) intervienen en el
procesamiento de los ARNr.
Se han descrito alrededor de 200 RNAsno. Estos
ARNsno se unen a proteínas y da una partícula ribonucleoproteica denominada
RNPsno. Similar a lo que sucede con el ARNm.
Las subunidades son sólo fucnionales cuando pasan el complejo
de poro y pasan al citoplasma.
Dentro del nucleolo se distinguen distintas porciones que son
3:
- Centro
fibrilar: son las zonas del nucleolo que son más claras y se corresponden con
el lugar donde se encuentran los genes del ARNr. Ese lugar estaría ocupado por
fibras de ADN.
- Componente
fibrilar denso: son las zonas alrededor de los centros fibrilares que se ven
más densas a los electrones. Y esas zonas se corresponden con los lugares donde
hay pre-ARNr.
- Componente granular:
son zonas donde se encuentran ya las partículas pre-ribosómicas. Es donde se
une el ARNr a las proteínas.
El nucleolo se desorganiza durante la división
celular. Y al final de la división el nucleolo se vuelve a organizar y se
organiza entorno a las regiones de los organizadores nucleares
No hay comentarios:
Publicar un comentario
TU COMENTARIO O SUGERENCIA NOS HARÁ CRECER